







作者:Jinwen 生信人|微信公众号
The roles of circRNAs in human cancer
人类癌症与环状 RNA
Publish: Ruoyu, Z., et.al., 2018, Cancer Letter
Translator: jinwen
简述
近几年关于 circRNA 的研究如火如荼,当前,我们也已经对 circRNA 有个大概的认识,比如什么是 circRNA、它是怎么形成的以及它在不同物种不同组织中普遍存在。据 circBank 的不完全统计,人类细胞和组织中大约发现了 14 万条 circRNA 转录本。这么多的 circRNA 到底具有什么样的功能呢,其实最终的问题是它对我们人类有什么用呢?有钱且疯狂的中国研究者不惜将人类所有疾病都测序一遍,最后发现 circRNA 分子确实在疾病中差异表达了。有差异表达了,好嘛,可以讲故事了……
今天要讲的故事是关于人类癌症与 circRNA 之间在近几年发生的那些事儿,主要包含以下几个方面:
circRNA 调控亲本基因(parental gene or host gene)
circRNA 充当 miRNA 的海绵
circRNA 与蛋白质形成功能复合物
circRNA 能翻译成蛋白质
而 circRNA 与癌症到底有什么关系呢,关于癌症的十个 hallmarks 大家都已经耳熟能详了吧,如果还不十分清楚,可以再回顾一下。而 circRNA 呢,却能影响癌症细胞的增殖、浸润与转移、血管生成以及细胞凋亡。
研究者最关心的还是 circRNA 在临床上到底有什么应用:
circRNA 不会让大家失望,研究发现 circRNA 与肿瘤 TNM 分型密切相关;除此之外,circRNA 能够在外泌体与体液中检测到,即能通过外周血检测 circRNA;另外,circRNA 的 RT-PCR 以及原位杂交比蛋白质更敏感和特异。这些都说明了 circRNA 作为肿瘤的生物标志物很有前景。
同时,circRNA 通过调控大量信号通路与肿瘤细胞的增殖、凋亡以及转移密切关联,另外还能通过细胞外囊泡(e.g. 外泌体,纳米微粒)进行转运,同时circRNA 具有低分子量、较好的稳定性与保守性的特点,因此,circRNA 具有作为治疗肿瘤的药物靶点或载体的潜能。
下表列出了与人类癌症关联的 circRNA 分子:
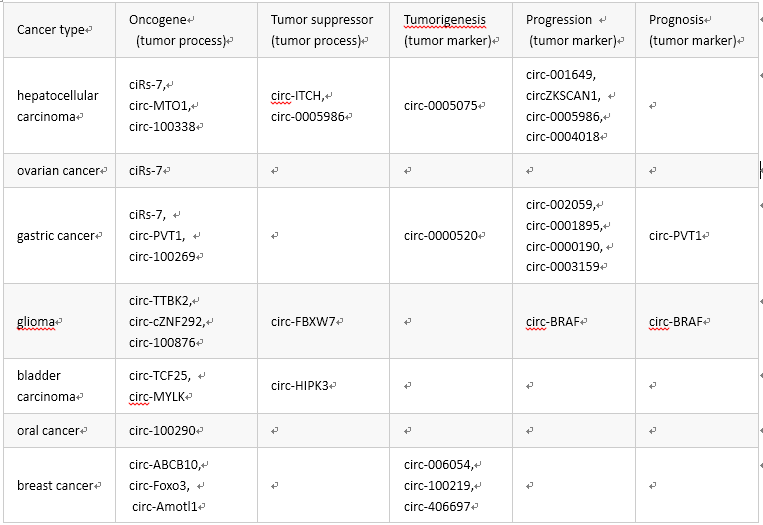
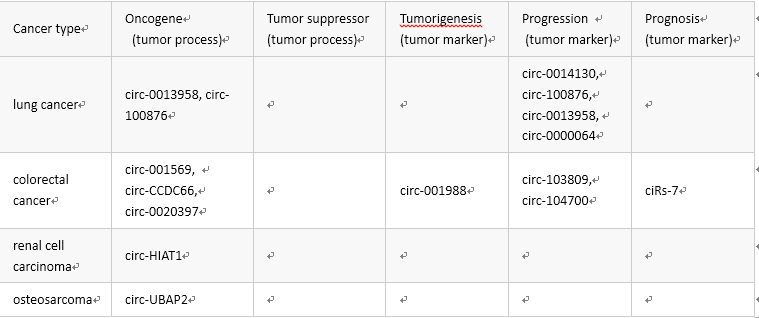
当然,做研究的都知道,梦想和现实往往是“很”有差距的,临床转化具有很大的挑战。当然,一切都是概率事件,只要概率不等于 0 那就是有可能;就像 2018 年的俄罗斯世界杯,谁能想到德国爆冷首先出局呢?所以,我们首先夯实理论基础吧。
circRNA 功能
circRNA 调控亲本基因
目前的研究表明,circRNA 是由线性 RNA 前体(linear RNA precursor)通过剪切加工形成的。其中,circRNA 可能由一小段 pre-mRNA 环化形成,因此,有研究者表明有一部分 circRNA 是 mRNA 的共转录产物。关于目前 circRNA 的环化机制可以用一张图总结,我们可以发现形成过程中,侧翼序列元件非常关键(e.g. ALU 重复元件):
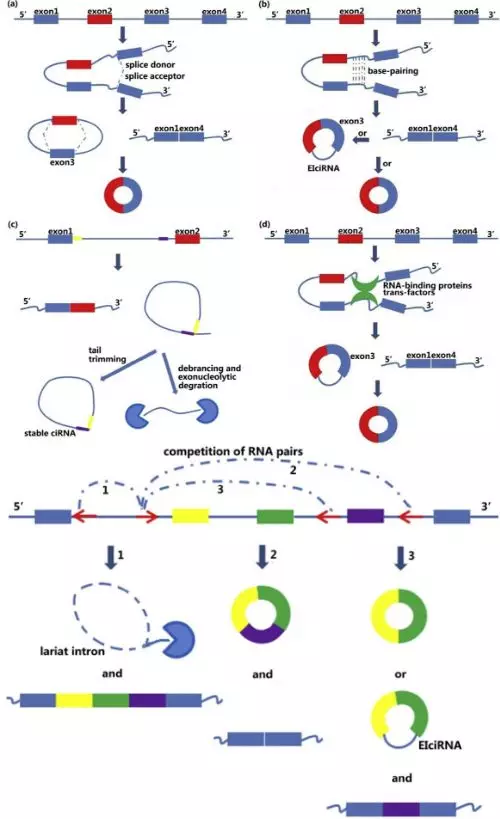
另外,有研究表明,一些 circRNA 分子能够在转录和转录后对亲本基因进行调控。
转录调控
EIciRNA 位于细胞核内,能够作为转录调控子。
Li et.al. 发现 EIciRNA 能够以 RNA-RNA 互作的方式与 U1 snRNP 结合,其中U1 与 RNA 聚合酶 II 形成复合物结合到 TFIIH 基因上。EIciRNA 通过调控 RNA 聚合酶II 的活性促进亲本基因的表达
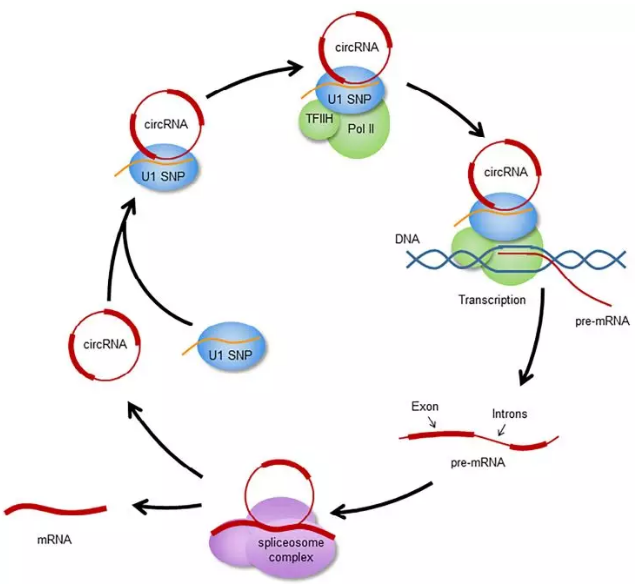
相似地,原核细胞转录起始因子(translation initiation factor eIF) 以及多聚腺苷结合蛋白(poly(A)-binding protein, PABP)的互作蛋白(PAIP)也是通过上面的机制增强表达的。
其中,eIF 在许多肿瘤中差异表达,例如肝癌、结肠癌,这表明,circRNA 的表达与肿瘤表型密切相关。
而 PABP 蛋白质与肿瘤的浸润和转移密切相关,PAIP 能够通过与 PABP 互作发挥相似的功能影响肿瘤的发展。
另外,ciRNA 也被发现定位在核内,并且上调 RNA 聚合酶II 的活性促进亲本基因的转录<=。
实验表明,ciRNA 能增强锚蛋白重复序列蛋白质(Ankyrin Repeat Domain 52, ANKRD52)的表达。
另外,circRNA 也能结合 U2,但这种关系目前并没有更进一步的研究。
snRNA
small nuclear RNA
小核 RNA 是真核生物转录加工过程中 RNA 剪接体的主要成分。
其长度在哺乳动物中约为100-215个核苷酸,共分为7类,由于含U丰富,故编号为U1~U7。
转录后调控
许多种类的 RNA 结合蛋白(RBP),包括 HuR(human antigen R)、AGO(argonaute)以及 TTP(Tristetraprolin) 都参与了 RNA 的剪切、稳定性调控以及 mRNA 的翻译,与细胞增殖、分化、运动、衰老以及凋亡密切相关。有研究预测 circRNA 分子序列上包含了特定的 RBP 结合位点,与 RBP 结合形成 RNA-protein 复合物(RPC)从而抑制特定 RBP 的功能,从而抑制亲本基因的转录。
Abdelmohsen, K., et.al. 就发现 circPABPN1 结合到了 HuR 从而抑制了 HuR 结合到 PABPN1 mRNA,导致 PABPN1(polyadenylate-binding nuclear protein 1)的翻译减少;同时,PABPN1 的减少会使 poly(A) 尾的形成中断,这样,正常生理过程中所需要的蛋白质被截短,最终造成代谢紊乱——这一过程也与肿瘤发生密切相关。
其他机制
circRNA 在转录及转录后调控亲本基因的机制有很多,但目前研究比较清楚还是结合 RNA 聚合酶II 以及 RBP。
除此之外,circRNA 能够以与 DNA 杂交或形成 R 环的形式增加同源外显子不足 mRNA(homologous exon-deficient mRNA) 的裂解效率,从而暂停转录并增加剪切因子<=。
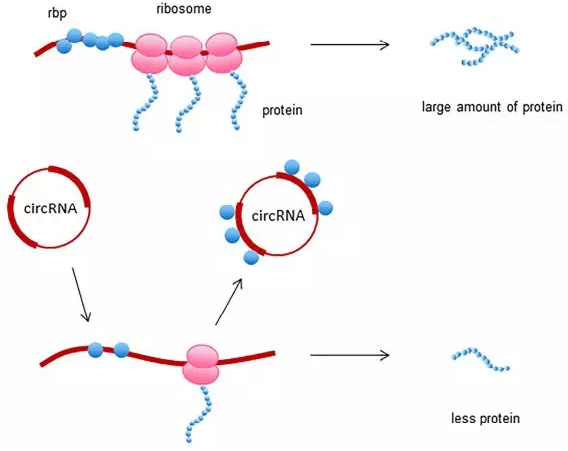
circRNA 充当 miRNA 的海绵
在细胞质中,circRNA 序列上也包含 MREs(miRNA response elements)作为 ceRNA 充当海绵吸附 miRNA,从而调控靶基因的丰度。circRNA 由于没有 poly(A) 尾和 5‘ 末端,较之线性 RNA 分子更不易被降解,具有作为 miRNA 海绵的优势。目前越来越多的研究表明,circRNA 能够发挥 miRNA 海绵的功能并且参与肿瘤的增殖、浸润、转移、凋亡以及血管生成。
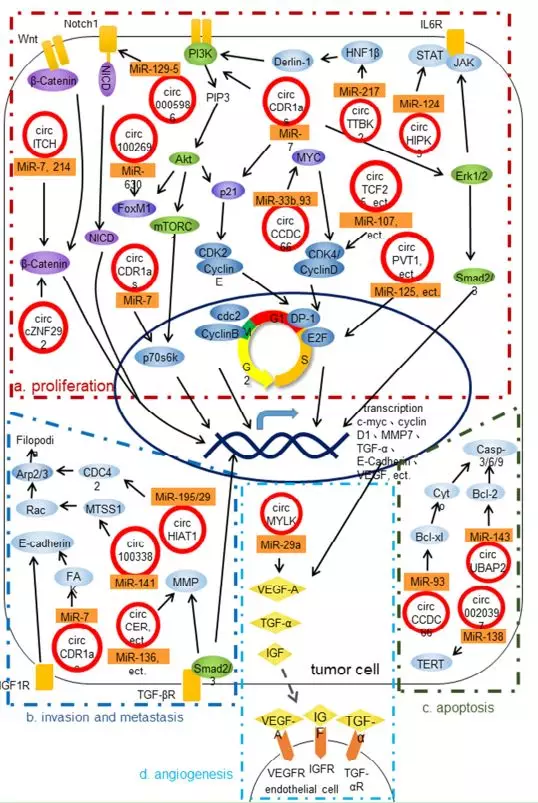
图中总结了 circRNA 对肿瘤相关 hallmarks 以及 基因的影响,而下面是详细的阐述
对增殖的影响
肿瘤细胞一个主要的特征就是过度增殖,而许多研究表明,circRNA 能够以作为 miRNA 海绵的方式通过调控细胞周期来影响细胞增殖过程。
肿瘤增殖被多个细胞通路所调控,而一些 circRNA 参与了这些通路的调控:
PI3K/AKT/mTOR 信号通路能够通过激活 CDK(cyclin-dependent protein kinase) 促进细胞周期的转变,其中 CDK 过度激活是肿瘤细胞增殖失控的首要原因<=。
cirs-7 能吸附 miR-7 导致 PIK3CD,p70S6K,mTOR 基因的上调,从而增大了肿瘤细胞增殖与转移的比率。
circ-TTBK2 通过与 miR-217 能够抑制肿瘤的发生。胶质瘤细胞中,miR-217 能够上调 HNF1beta(hepatocyte nuclear factor1 beta)的表达,从而激活 Derlin-1 并通过 PI3K/AKT,ERK 及其他通路促进肿瘤增殖、浸润以及转移。
MAPK/ERK 信号通路也与肿瘤细胞增殖密切相关。ERK(extracellular regulate protein kinase) 蛋白质被蛋白激酶 Raf 激活,从而充当多种转录因子通过级联反应调控下游癌症相关基因的表达。
miR-7 能够抑制 ERK1/2 的活性,而 cirs-7 作为海绵可以激活 MAPK/ERK 通路从而促进细胞增殖<=。
Wnt/β-Catenin 信号通路中,β-Catenin 能够进入细胞核激活下游靶基因 c-myc, cyclin D1, mmp7, E-Cadherin, VEGF, etc.的转录,而这些基因能够促进肿瘤的发生过程。
在胶质瘤细胞中沉默 circRNA-cZNF292 将会降低转录因子(β-catenin, p-STAT3, p-STAT5)以及下游分子(cyclin A, CDK2)的表达,也就是说 cZNF292 可以通过 Wnt/β-Catenin 信号通路影响肿瘤的增殖,然而其中的机制目前仍不清楚<=。
另外,circ-ITCH 能够结合 miR-7 以及 miR-214,从而通过下调 β-Catenin 抑制肿瘤的增殖<=。
Notch 信号通路也参与了肿瘤的发生发展。
Hsa_circ_0005986 在肝细胞癌(HCC)中的减少使 miR-129-5 表达增加,从而抑制了 Notch1 的活性,最终促进了细胞周期从 G0/G1 向 S 期转变。
细胞周期相关蛋白或转录因子也能被 circRNA 通过 miRNA 简介影响,最终调控肿瘤的增殖。
circ-TCF29以及 circRNA-100290能够 miRNA 结合(e. g miR-107, miR-103a-3p)增强 CDK6 的功能。
circ-ABCB10与 hsa-circ-0013958可以分别作用于 miR-1271 与 miR-134 从而增强 CDK1的功能以及上调 cyclin D1 的表达,最终促进肿瘤增殖。
E2F 转录因子家族在细胞周期 G1 到 S 期过程中扮演着重要角色,许多 circRNAs 都参与调控这些蛋白质。例如,circPVT1 能竞争性结合 miR-125 家族分子从而增强 E2F 蛋白质的表达(Jie Chen et.al.);circRNA-001569 与 miR-145 互作增强 E2F 家族的表达(HuijunXie)。
与细胞增殖相关的 circRNA 与通路还包括 :circRNA-CCD66 能作用于 miR-33b 和 miR-93 从而增强了myc 与促进细胞增殖的基因的结合<=;circHIPK3 作为 miR-124 的海绵在一定程度上能够反转 miR-124 对靶基因 IL6R(interleukin 6 receptor) 和 DLX2(Distal-Less Homeobox2) 的抑制作用,最终抑制细胞增殖<=。
对浸润以及转移的影响
肿瘤细胞不仅能够浸润临近组织,还能够转移到远端的器官从而形成新的肿瘤。在这写过程中,细胞会发生一些改变,例如过度增殖、上皮间充质转换(EMT)以及基质金属蛋白酶过表达(MMP)。circRNA 参与了癌细胞的浸润与转移的过程。
PI3K/Akt信号通路是癌细胞浸润及转移最重要的通路之一,PI3K(phosphoinositide 3 kinase) 的激活能生成PIP3(phosphatidylinositol),随后会激活 Akt(serine-threonine kinase),从而促进下游靶基因的转录,同时调控细胞浸润与转移。除此之外,PIP3 能够激活 cdc42,这是一种 Rho 蛋白质——能够直接作用于激动蛋白调控蛋白(例如 Arp2/3 复合物),从而促进丝状伪足<= 的形成并减少粘着分子(例如 E-cadherin)<=。
在几种癌症中已报道<=,circRNA-7 能够吸附 miR-7 从而阻止 PI3K/Akt 通路,并通过调控 IGF1R(Insulin Like Growth Factor 1 Receptor) 与 FAK(Focal Adhesion Kinase) 增加粘着分子 E-cadherin 的含量。最终 circRNA-7 改变了癌症的浸润与转移。
另外,circRNA_100269 具有相似的作用<=,它在胃癌细胞系中与 miR-630 表达负相关,而 miR-630 能够通过调控 PI3K/Akt 信号通路中的 FoxM1 蛋白质抑制 EMT 过程。
另一个关于肾细胞癌<=表明 circRNA HIAT1 能够抑制 miR-195/29 的浓度,从而激活 cdc42,最终刺激丝状伪足的形成并调控粘着分子的表达。
靶向 MMP 的 miRNA 可以被 circRNA 直接抑制,从而影响肿瘤细胞的浸润与转移。
在非小细胞肺癌中,circRNA_100876 能够作为 miR-136 的海绵,其中 miR-136 能够抑制 MMP-13 mRNA 的表达水平[](42。
同样地,肝细胞癌中 circRNA-MTO1 降低了 miR-9 的浓度从而调节 MMP-14 的分子水平。
此外,circRNA_100338 被发现可以作为 miRNA-141 的海绵,而该 miRNA 能够抑制肿瘤抑制基因 MTSS1 的表达。
对血管生成的影响
血管生成是肿瘤十个基本特征之一,在肿瘤发展过程中起到了关键作用。“血管生成的开关“,包括 TGF(transforming growth factor)、VEGF(vascular endothelial growth factor) 和 MMPS(matrix metalloproteinases),与血管生成的调控紧密关联。而 circRNA 却能够通过介导 miRNA 来调控这些因子的表达,最终调控肿瘤的血管生成。
circ-100876(circ-CER) 能够抑制 miR-136 与 MMP-13 结合,从而上调 MMP-13 的表达<=;相反却影响 ALK-1(Actin receptor-like kinase 1)与 ALK-5 的关系从而增加 VEGF 的丰度<=,最终促进血管生成。
circ-MYLK 通过吸附 miR-29a 影响 VEGF-A 的表达<=。
对细胞凋亡的影响
细胞凋亡对于维持细胞内稳定非常重要,然而肿瘤细胞往往能够逃避凋亡,使发生异常的细胞不能被清除。细胞凋亡的触发被 Bcl-2 家族的蛋白质所控制着,同时这些蛋白质也能够调控肿瘤细胞的 caspase 信号通路<=。
circRNA UBAP2 与 miR-143 结合上调了 Bcl-2 的表达,从而阻断了 caspase 凋亡通路<=。
circ-CCDC66 被发现能够增加抑凋亡因子 bcl-xl 的表达<=。
另外,端粒酶异常表达能够降低对肿瘤细胞增殖的限制,这也是肿瘤细胞能够抑制凋亡的主要原因之一<=。
circ_0020397 能够减少 miR-138 的浓度,从而增加端粒酶的表达水平,从而抑制了肿瘤细胞的凋亡<=。
circRNA 与蛋白质形成功能复合物
circRNA 能够与蛋白质形成复合物,在细胞胞中发挥重要的功能。
circ-Foxo3 在非癌细胞中高表达,而该 circRNA 的异常表达会抑制细胞增殖——其中 circ-Foxo3 与细胞周期蛋白质 CDK2(cyclin-dependent kinase 2) 以及 p21(cyclin-dependent kinase inbitor) 结合,而异常的 circ-Foxo3 使 p21 更容易抑制 CDK2;相应地,CDK2 无法与 cyclin A 或 cyclin C 互作,从而使细胞周期停滞在了 G1期,阻断了细胞发展的进程。
circ-Foxo3 与 MDM2(Mouse double minute 2 homolog) 以及 p53 能形成三元复合物,该复合物通过促进 MDM2 介导的泛素化以及降解减少 p53 的丰度。另外, MDM2街道的 Foxo3 泛素化被下调。最终,下游 Foxo3 靶分子 PUMA(P53 Up-Regulated Modulator of Apoptosis) 表达上调从而降低了细胞凋亡<=。
除此之外,circ-Foxo3 可以与 CDK6,p16,p27 结合,表明它能够通过其他通路影响细胞周期。
circ-Amotl1 被发现能够通过 c-myc 促进增殖并抑制凋亡,研究表明 circ-Amotl1 能够与 c-myc 共定位并且互作。circ-Amotl1 促进了 c-myc 在核内的滞留,增强了 c-myc 的稳定性并上调了它的靶基因。Amotl1 的表达也能增加 c-myc 结合到启动子的亲和力,最终导致肿瘤形成。这一过程展现了 circRNA 与蛋白质结合的新机制,表明 circRNA 能够促进癌症的进展<=。
circRNA 翻译成蛋白质
circRNA 能翻译蛋白质可查阅的历史要追溯到 1984 年,Wang ZeFeng 等人发现 circRNA 可以由一条人工设计的内部核糖体进入位点(IRES,internal ribosomal entry site)产生功能性的绿色荧光蛋白(GFP, green fluorescent protein)。
Abe, H 等人发现 circRNA 包含的开放阅读框(ORF)可以被有效的翻译成蛋白质。
现在的研究表明,即使没有 IRES 、poly(A)尾 或 5' 帽结构,翻译也能够进行。
目前已被证实能够翻译蛋白质的 circRNA 包括
circ-Mbl
circ-ZNF609
circ-FBXW7
该 circRNA 被发现能够编码一个 21-kDa 的蛋白质 FBXW7-185aa,在癌症细胞中上调该蛋白质将会缩短 c-Myc 的半衰期,从而抑制增殖并加速细胞周期。
目前预测 circRNA 编码能力的数据库包括:
circRNADb

来第一个抢占沙发评论吧!