







模式植物水稻和拟南芥转录组研究显示环状RNA广泛分布于植物中,同时植物环状RNA与动物一样存在特定的时空表达模式;此外,植物环状RNA与动物相比具有一些明显的特征,如非non-GT/AG剪切信号的环状RNA大量存在。
对于农学经济植物的研究也是近两年才刚刚起步(表1),目前主要工作集中于环状RNA数量鉴定和差异表达研究,对植物环状RNA的功能作用和形成机制等深入研究还存在很大的空白。
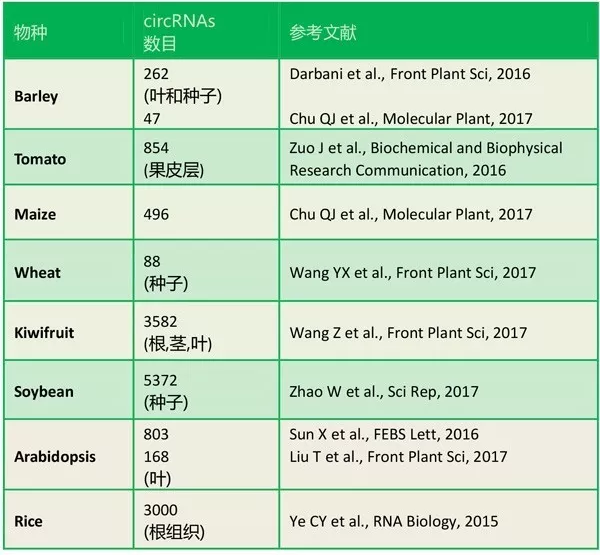
表1. 已报道的植物circRNA
下面以一篇文章为例,看看植物环状RNA目前的主要研究内容,以及WGCNA在其中的应用。
研究背景
猕猴桃(Kiwifruit)是一种重要的经济水果,Pseudomonas syringae pv. actinidiae (Psa)引起的溃烂病(canker disease)给全球的产业带来严重的损失。由于对宿主−病原互作相关分子生物学机制了解很少,目前多种病害控制策略都不能很好的对病原(Psa)感染进行有效控制。尽管已有报道模式识别受体蛋白(PRRs)genes、resistance(R)genes和转录因子(TFs)与植物免疫有关,CircRNA和免疫基因间的关系以及在植物免疫应答中的作用还未报道过。
样本选择和建库
研究组织表达模式:样本Ac-derived cultivar “Hongyang”(AH)的叶、根和干组织(每个样本两个生物学重复,共6个去核糖体RNA文库)。
研究不同栽培种和品种中特有的、与病原感染相关circRNA:Psa感染A. chinensis(AH和AJ)、A. eriantha(Ae)和A. argute(Aa)叶组织后不同时间点day post incubation(0、2、14 DPI)(每个样本两个生物学重复,共24个去核糖体RNA文库)。
研究思路
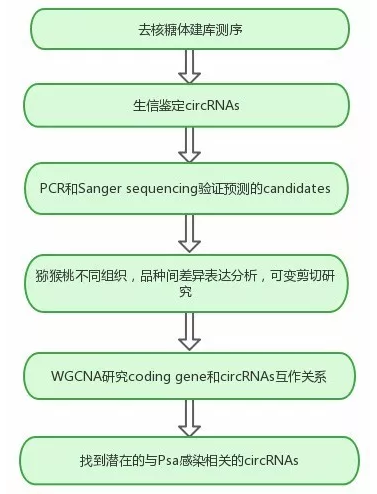
研究结果
1、circRNAs测序数量统计和可变剪切分析
在30个RNA文库中总共鉴定到3582个circRNAs,根据其基因组位置信息可分为:基因间区、外显子型和内含子型circRNAs。与水稻和拟南芥circRNAs主要来源于编码基因情况不同,猕猴桃不同组织和品种中基因间区circRNAs数量最多,约占51%和67%(图1A)。
此外,同一基因组区域可以通过可变剪切产生不同类型的circRNA isoforms。作者总共发现4种不同类型,总共163个可变剪切(图1B)。不同品种在感染不同时间点的可变剪切事件比较稳定,此外2 DPI和14 DPI的剪切数相同,暗示circRNA在感染过程中稳定表达(图1C)。随机选取80个circRNAs进行PCR验证,其中68个circRNAs能够被实验证实,并且不同类型circRNAs比例相同,说明不同类型的circRNA都稳定的表达(图1D)。
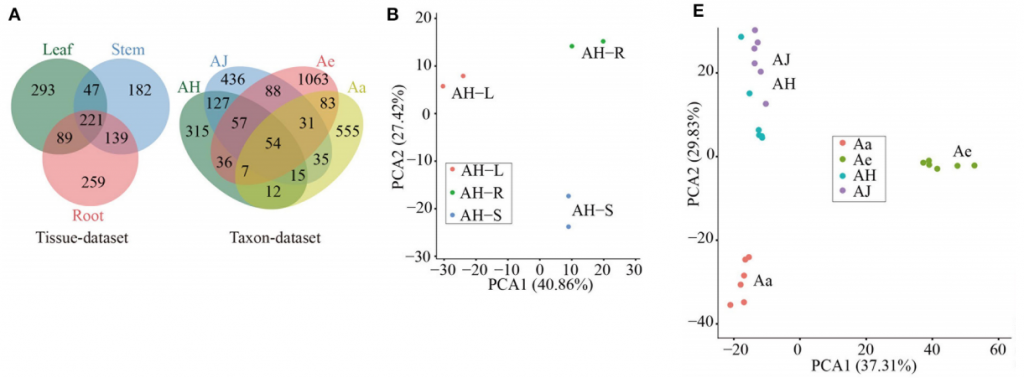
图1. (A) 不同猕猴桃组织和品种中三种类型circRNA数量分布
(B) 可变剪切发生情况(AS events):A3SS, alternative 3’ splice site;A5SS, alternative 5’ splice site;ER, exon skipping;IR, intron retention
(C) 病原感染不同栽培种和品种不同时间点可变剪切情况
(D) PCR和sanger测序验证Ac_ciRNA_11553示意图
2、猴桃circRNAs表达模式研究
对不同组织的circRNAs表达发现,约有59.67%只在一种组织中特异表达,三种组织中共有表达的只占17.97%。PCA也将三个不同组织聚成明显的三类,说明猕猴桃circRNAs具有组织特异性表达特征(图2A, 2B)。
不同栽培种/品种叶组织中,81.3%的circRNAs具有品种特异性表达特性(图2A),PCA也将样本聚成三类A. chinensis(AH和AJ)、A. eriantha(Ae)和A. argute(Aa)(图2E)。
3、猕猴桃circRNAs和来源基因的表达关系
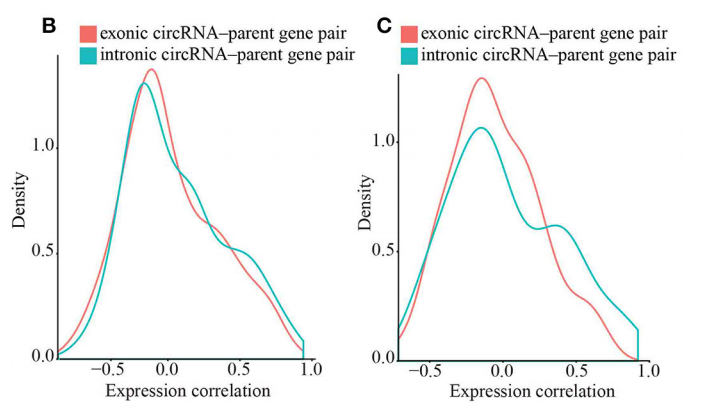
作者计算circRNAs和其来源基因的表达相关系数,发现exon-circRNAs和intron-circRNAs与来源基因分别有15.33%和19.64呈正相关,有8.33和5.19%与来源基因负相关(p < 0.05)。说明多数exon-circRNAs和intron-circRNAs能够提高其来源基因的表达(图3B)。
利用同时能够产生exon-circRNAs和intron-circRNAs的来源基因,比较exon-circRNAs和intron-circRNAs对其的影响,发现intron型的相关性比exon型要显著,说明intron-circRNAs在其来源基因的表达调控中扮演更重要的作用(图3C)。
4、circRNAs和编码基因共表达网络研究
为了研究circRNAs在病原感染过程中的作用,作者利用WGCNA挖掘相关基因。利用鉴定的circRNA数据和通过转录组数据注释的编码基因数据,先筛选品种间至少一个差异对的8700个编码基因和584个circRNAs,再将这些编码基因的FPKM matrix和circRNAs的RPM matrix整合一起进行共表达分析,总共得到28个相同表达模式的模块(图4)。
根据模块内基因将模块分为三大类,进而将模块基因与三种生物学特征关联起来,发现模块17和18显著与多种特征相关。28个模块中有15个与Trait 3(不同种)相关,作者重点关注这些种间差异的模块基因。
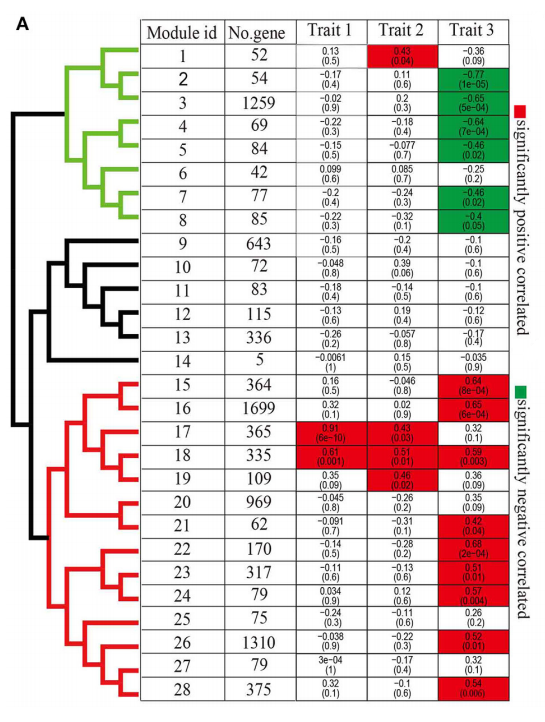
图4. WGCNA分析猕猴桃编码基因和circRNAs共表达模式
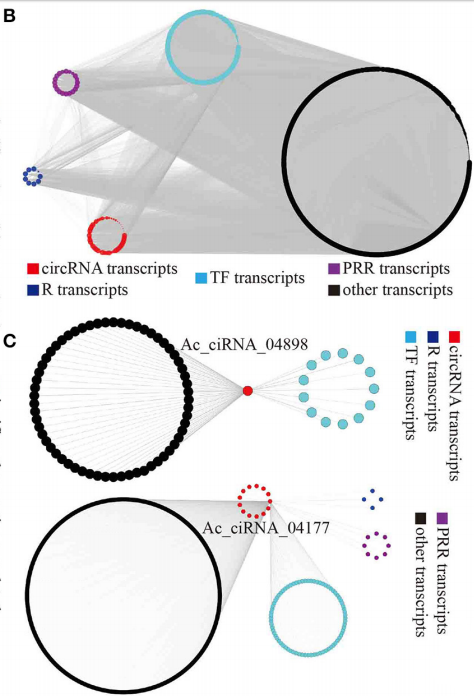
为了研究各品种间的病原易感和耐受情况,作者对种间显著相关的模块基因进行共表达分析,通过Cytoscape产生几个大的基因集包括:the R genes、PRR genes、TFs以及co-expressed circRNAs(图5B)。
根据WGCNA的结果,作者建立了一些subnetworks,主要关注特定circRNA与其相关的编码基因和circRNAs的关系,挖掘一批与circRNA相关的特定编码基因(CAPC)。利用GO对这些CAPC进行功能注释,发现显著关联于植物-病原互作功能,尤其在那些品种间差异的circRNA的网络中,这一结果暗示circRNA在植物-病原互作中发挥一定的功能(图5C)。
总结
作者设计了多样本测序策略,从多角度研究了猕猴桃不同栽培种/物种、不同组织中以及病原感染不同时间点的circRNA数量和表达模式,发现了circRNA具有组织特异性和种间特异性表达的特征。
通过研究circRNA和其来源基因的表达关系发现,正相关基因pairs多于负相关,说明猕猴桃circRNA对其来源基因表达有一定的促进作用。
利用WGCNA分析了多组编码基因和circRNAs和数据,将不同的模块基因与三种生物学特征关联,进而关注不同种间显著相关的模块基因,并对其功能进行注释,发现circRNAs参与了植物-病原互作的生物学过程。
这篇文章侧重于circRNA的表达模式研究,结果表明circRNA的生物学调控可能是比较复杂的,不仅本身具有特异性表达特征,还能通过编码基因-circRNA、circRNA-circRNA互作发挥功能。
对筛选的circRNAs的功能机制还需要做深入的研究,才能更好阐释植物circRNAs采用何种形式发挥功能,如:miRNA sponge、转录调控、蛋白翻译等。
参考文献:【1】Wang Z, Liu Y, Li D, et al. Identification of Circular RNAs in Kiwifruit and Their Species-Specific Response to Bacterial Canker Pathogen Invasion[J]. Frontiers in Plant Science, 2017, 8.
来源:基迪奥生物

来第一个抢占沙发评论吧!