







circular RNAs (circRNAs)是最近几年发现的一类内源共价闭合的环状RNAs。它们最初是1970s在植物类病毒中发现。但是随后的几十年,只有很少数的circRNAs被发现,由于cicRNAs的低表达量和组织表达特异性以及分子生物学技术的限制,最初它们被认为是转录的副产物或不正常剪切转录的噪音。但随着高通量测序技术和生物信息学分析技术的发展,成千上万的circRNAs被鉴定出来。它们在真核生物中广泛存在,并在多个生命活动过程和发育过程中发挥非常重要的功能。环状RNA与疾病的发生特别是肿瘤的发生和发展密切相关,已经成为RNA领域一个新的研究热点。
大部分circRNAs的研究是发生在动物中, 最近三、四年植物中的circRNAs才被报道。通过转录组水平的研究在多种植物中发现了大量circRNAs,对一些植物中circRNAs的产生和作用机制也有了初步研究。
植物circRNAs的鉴定
circRNAs可以从外显子,内含子以及基因间区产生。是在mRNA前体(pre-mRNA)剪切过程中,外显子的5’端和3’端以反向剪切方式形成的环状RNA,如图1(a)所示。生物信息学手段鉴定circRNAs的方法原理是根据动物中circRNAs的研究,分析去除核糖体RNA的全转录组数据库。首先去除可以比对到参考基因组上的reads,对那些不能比对到参考基因组上的reads,做进一步的分析。将这些reads的两端各取20-nt的reads称为anchor 序列,记为5’anchor和3’anchor,将每一对anchor序列再比对到参考基因组上,如果两个anchor 序列按相反的位置前后均能比对到参考基因组(5’anchor比对在右端,3’anchor比对在左端),并且该anchor所在的完整reads能在比对的位置找到剪接位点,并且完美匹配,则将此完整的reads作为候选的circRNAs[图1(b)]。最后再进行某些条件的筛选。目前常用的用于circRNAs鉴定的工具有MapSplice,PredcircRNA, find_circ_enhance等等。
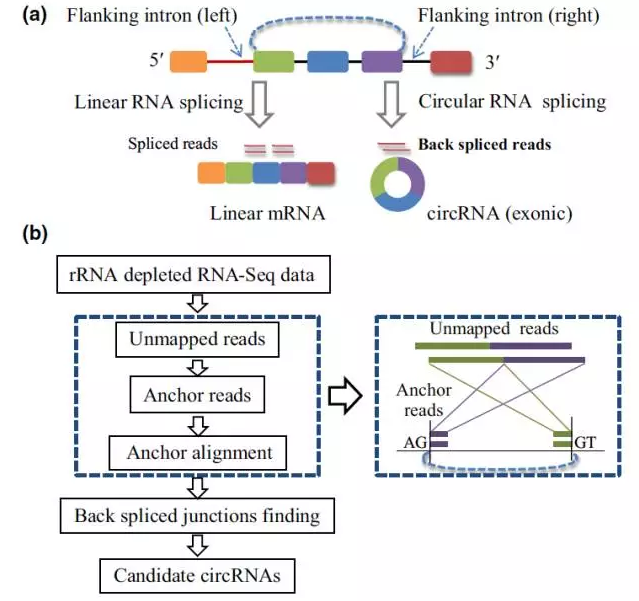
目前在拟南芥叶片和水稻根中鉴定出6012和 12037个circRNAs,而且至少有700个分子是同源的,说明circRNAs在不同植物中存在着保守性。在番茄、小麦、大麦、猕猴桃等植物中也鉴定出了大量circRNAs,揭示出circRNAs在植物不同物种中是广泛存在的。虽然我们已经发现了很多植物中的circRNAs,但是真实的数量远远不止这些。因为目前大部分研究植物circRNAs的方法都是来源于人类或其它动物的研究,这就可能造成忽视植物基因组的特性和植物circRNAs产生方式的不同。目前的研究表明,在动物中环状RNA外显子两侧翼的内含子中有大量的ALU和其反向互补重复序列,它们可以通过和内含子的配对竞争来调控外显子的环化。但是在植物中很少有这样的重复序列。对于水稻circRNAs的研究表明,只有10%的circRNAs有典型的GT/AG (CT/AC)剪切信号,大部分的circRNAs的两端是非GT/AG。这就和动物中大部分是典型GT/AG不同。最近出现的PcircRNA_finder工具可以专门针对植物circRNAs进行研究,为未来发现更多植物来源的circRNAs提供了重要的支持。
植物circRNA的作用机制
a)miRNA 海绵(sponge) 可能不是植物circRNAs的主要功能
目前有些研究表明circRNAs可以作为miRNA海绵,结合miRNAs来上调miRNA靶基因的表达 (图2)。而只有很少量的circRNAs满足作为miRNAs海绵的条件,这就说明作为miRNA海绵可能不是circRNAs的主要功能。
植物中miRNA起作用的主要方式是依靠与其靶基因序列互补来切割靶基因。miRNAs和其靶基因在第9位到11位的完全互补对于miRNAs有效切割靶基因是必须的。因此预测miRNA靶基因类似物 (mimics),需要在第9位到12位之间有3个碱基的凸出。但是这个凸出会降低miRNA和其靶基因类似物的亲和性,减少了靶基因类似物的竞争力。要作为miRNA靶基因的类似物,就必须有足够多的miRNA结合位点或者高表达水平的circRNAs来补偿低亲和性的影响。但是目前发现的植物circRNAs都没有大量的miRNA结合位点,而且circRNAs的表达量都很低。在拟南芥中只有5%的circRNAs被预测是miRNAs靶基因类似物。
circRNAs二级结构可能会隐藏部分的miRNA结合位点,而在预测的时候也可能会预测过多miRNA结合位点。随着越来越多的circRNAs被发现,对于circRNAs作为miRNA靶基因的功能会引起更多的重视。
b)植物circRNAs在应答生物和非生物胁迫过程中的作用
有研究表明circRNAs在各种胁迫条件下表达会发生变化。比如,在水稻低磷和多磷胁迫下有27个外显子circRNAs表达发生变化。在干旱胁迫下小麦中62个circRNAs表达发生变化。猕猴桃中的研究发现circRNAs对于病原的入侵有特异的应答反应。这些都说明了circRNAs参与了应答生物和非生物胁迫。
circRNAs有一个封闭的环状结构,没有自由的5’和3’端,也没有poly(A)尾,因比线性RNAs更稳定,不容易被RNA核酸外切酶降解。某些circRNAs可能因为稳定性,可以作为应答生物和非生物胁迫的慢速调控因子。同时在动物中的研究发现circRNAs可以在外泌体中富集和稳定。外泌体是一个小的囊泡结构,包含很多蛋白、mRNA和非编码RNAs,它们在某些特定细胞中可以被释放到细胞外的微环境中,调控细胞的生命行为。
因此,植物circRNAs可能作为信号分子参与细胞之间信号传递和木质部韧皮部的长距离运输。第一个发现的circRNA是来源于植物类病毒,这些类病毒circRNAs的不同结构域可以结合不同的RNA结合蛋白参与病毒细胞间和长距离的运输。推测植物内源的circRNAs可能采取类似的方式参与信号传递。
c)circRNAs调控亲本基因的表达
circRNAs的形成依赖于侧翼RNA元件。侧翼RNA元件可能对内含子环化非常重要。研究发现一些circRNAs定位在细胞核中,可以与通过cis或trans作用模式调控宿主的转录活性。拟南芥的一个来源于SEP3基因6号外显子的circRNA可以结合到SEP3基因形成R-loop造成转录暂时停滞,而导致可变剪切过程跳过该外显子,产生缺少该外显子的转录本从而降低亲本基因的表达。circRNAs的功能也和其亚细胞定位密切相关,对于circRNAs亚细胞定位的研究会对揭示circRNAs的生物学功能有重要意义。
另外, 有一小部分circRNAs是可以翻译成蛋白的。研究发现人工合成的在起始位点上游插入IRES的circRNAs可以翻译蛋白。同时由于circRNAs的环状结构它可以获得开放阅读框,而翻译为长的含有重复序列的蛋白。
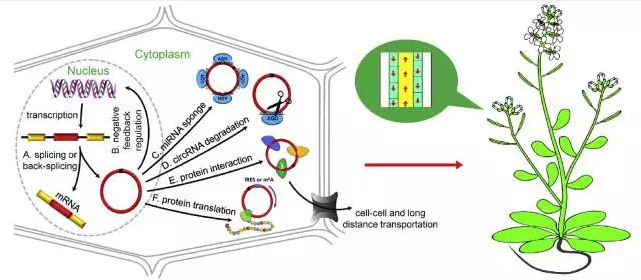
图2 植物中circRNAs的产生和调控机制
高通量测序技术和生物信息技术的迅速发展发现circRNAs在植物中广泛存在,而且有较强组织表达特异性和参与胁迫响应,揭示circRNAs在植物生长发育中有重要的作用。在动物中的研究已经揭示出circRNAs的多种重要功能和作用方式,是否植物circRNAs有着同样的分子机制需要更进一步的研究。另外一方面关于植物circRNAs的产生和降解机制亟待解决。circRNAs在其它植物种类中也需要进一步的研究。作为一个新的研究热点,虽然circRNAs在植物中已经取得了一些进展,但是得到清楚和深入的产生、功能、调控机制等还有很长的路要走。
首发:一呼百诺 |公众号ID:allfrombionova

来第一个抢占沙发评论吧!